Biology Worthy of Life
An experiment in revivifying biology
Who Are You and Who Am I and Who Are We?
Stephen L. Talbott
In every sphere of knowledge it’s easy to put out of mind those questions that are so fundamental and yet so seemingly impenetrable that they leave everything we think we understand woefully ungrounded. Biology is no exception.
Who, or what, is the organism, and what guarantees the remarkable unity of character, the distinctive and recognizable way of being, consistently achieved by the developing individuals of a species? Where do we look for the guarantor of this unity if we are unwilling to commit the obvious error of making a particular part of the organism — or an ever-varying collection of parts — an explanation for the integral performance of the whole?
And what is such a whole? Is every organism an unambiguous and definable unity, so that we can precisely delineate the boundary separating it from other organisms and from the larger environment? Surely this boundary becomes blurred in numerous ways, ranging from obligate symbiosis (where neither of two organisms can live except in intimate relation to the other) to natural cloning (where, for example, the quaking aspen trees covering entire hillsides may arise from a single, shared root system).
Or again: what do we mean when, regarding ourselves, we speak, as so many do, of a psychosomatic unity? Who will specify exactly what is meant by the psychological half of “psychosomatic”? And whatever is meant by it, how and in what sense do psyche and soma become a unity? — a question neither biologists nor cognitive scientists nor philosophers have been able to resolve with any hint of consensus, despite centuries of effort. Looking beyond ourselves, in what sense do we find anything like a psychosomatic unity in chimpanzee, nightingale, cricket, or amoeba? If we cannot answer such a basic question despite our being driven to employ a descriptive language that includes both physical and psychic terminology, then what do we understand?
Two problematic themes running through these questions, as we will see, are those of identity and context-sensitivity, or wholeness. How, precisely, do we define a protein, when its form and function depend on the molecules with which it associates? How do we understand the identity of a cell in the developing embryo, when it can become any one of many different kinds of cell, depending on where it migrates to? And when we find the same plant with a radically different character in different environments, what are we recognizing as the “same”? How, if at all, can we conceive the separate identity of something that is always taking on the character of its larger context?
“Organism”, “species”, “individual”, and, more recently, “context” — these are fundamental, if taken-for-granted terms of biology, terms upon which large theoretical edifices have been erected. But how secure can those edifices be, if the foundation stones lack definition?
I wish to underscore the difficulty and importance of the foregoing questions, and also to make some suggestions about ways we will need to change our thinking in order to address them profitably. But first we need at least one example of research that gives concrete form to the questions.
Which comes first: the cell or its niche?
Every cell in an organism lives in a sufficiently distinctive way — is enough of a whole in its own right — to pose the question of identity. What makes this particular cell of my skin a keratinocyte rather than a melanocyte? Does identity imply constancy of cellular character? To what degree does a cell’s environment — the larger community of cells — shape identity and character?The December 19 issue of Cell contains a brief report on recent researches showing how the fate of stem cells is bound up with that of nearby cells in the same local environment, or “niche” (Tan 2013*). One of the studies mentioned in the article concerns the basal stem cells of the airway. (Basal cells are epithelial cells constituting the lowest layer of epidermis, and stem cells are relatively undifferentiated cells capable, at need, of dividing and differentiating into more specialized cell types.)
According to the cited study,
When airway basal stem cells are in demand, spontaneous reprogramming occurs in vivo as committed secretory cells convert into stem cells to replenish the stem cell pool. ... [The] secretory cells proliferate, acquire stem cell markers, and eventually generate a stable stem cell pool that gives rise to all three epithelial cell types of the airway.
Further, direct contact with a single basal stem cell is enough to prevent secretory cells from dedifferentiating and becoming stem cells. Clearly, then, the identity of these fully matured secretory cells is not rigidly fixed, and at the same time their transformation potential is delicately sensitive to context. We can hardly separate the question of a cell’s identity from that of the niche’s identity.
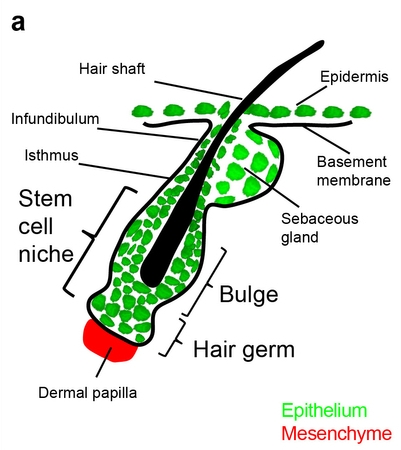
Mouse hair follicle. (From Rompolas et al. 2014*.)
This point is driven home by a second study concerning mouse hair follicles (Rompolas et al. 2014*. See illustration at right.) The researchers explored how a cell’s location within various compartments of the niche affects its fate. For example, stem cells in the bulge tend to stay quiescent — that is, they remain in a resting state without cell division — whereas those in the hair germ are continually differentiating into more specialized cell types. And even within the bulge, stem cells in the upper half remain much more consistently quiescent, whereas those in the lower half are more proliferative.
Dramatically, the authors also show that “niche stem cells can be dispensable for tissue regeneration, provided that the overall integrity of the niche is maintained”. When the stem cell population in the bulge or hair germ is destroyed by laser ablation, distant epithelial cells flow toward the damaged compartment and go through a transformation of identity enabling them to replace the lost cells. As the authors summarize it, “The overall structure and function of the tissue is maintained because cells are capable of adopting new fates as dictated by their new niche microenvironment”.
It is impossible to reconcile these goings-on in the hair follicle with the picture of an organism being constructed from an available collection of well-defined parts as building blocks. The larger context helps to “decide” what sorts of elements it will have, how they should be transformed, and how they will come into mutual relationship. Nothing could be further from the common picture of the organism or the cell as a product of bottom-up causation, where the sole basis for understanding consists of putting back together in our minds the parts we have previously analyzed out of — and severed from — their life-receiving connection to the whole.
This is the kind of finding that might well be shouted from the rooftops, because it turns longstanding habits of biological thinking upside down. Actually, it is being shouted — ten thousand times over1 — from the pages of technical journals. Context-dependence has become a devoutly repeated mantra in biology, from molecular and cell biology to evolution, as well as in a field such as ecology, where it was pretty much the whole point from the beginning.
Facing up to the questions
The author of that article in Cell ends her review of each piece of research on stem cell niche-dependency with a question. She wonders, for example, how “different niche cells at different locations are able to maintain different status”.This is a perfectly good question, deserving exploration. But much depends on how we approach the task, and the signs are not particularly hopeful when the question is accompanied by references to “the intrinsic signaling elements that may dictate the behavior” of certain cells, and to “the molecular switch controlling cell proliferation” (my emphasis). According to this kind of thinking, individual parts with stable functional identities have the kind of unqualified determining effects indicated by words such as “dictate” and “control”.
If these terms were justified, they would nullify the idea of context-sensitivity. But the whole point of the research under discussion is that cells do not have absolutely stable functional identities. The context as such is having its say. And even if biologists have so far spent precious little time considering what they mean by “context”, this does not justify pretending that their own words can be safely ignored.
The simultaneous use of the language of context and the language of the controlling part is ubiquitous in the literature today, despite the brute contradiction involved. The irony is that those who speak of controlling parts are generally inclined to ridicule notions of holism. And yet they are, in effect, turning their wonderfully autonomous causal parts into absolute and determinate wholes — the kind of context-independent wholes we never find anywhere in the organism.
Even a superficial acquaintance with the biological literature today makes it clear that the effort to explain the whole organism as a result of self-contained causal parts that do not participate in, and are not sustained by, the life of their larger context is destined to fail. If biologists are going to speak incessantly about the importance of context, then they need to reckon honestly with the problems it raises, rather than immediately change the subject to “controlling factors” — factors that, as the contemporary literature makes so vividly clear, never do control in absolute terms2. If all such factors are context-dependent, we ought at least to ask ourselves how this qualification modifies the notion of “control”.
We are, in fact, beset by questions on all sides. What exactly are we referring to when we speak of “context” and “organism”? How can we make these terms, as we are compelled to use them, more than necessary blanks in our descriptive language — blanks about whose necessity we can say nothing? For example, does our actual use of these terms differ much from the way people of an earlier era might have used “archetype” or “entelechy” or “being”?
When biologists speak of the organism’s activity, who exactly do they mean to say is performing that activity? When they acknowledge that something in the organism is context-dependent, what in fact is it dependent upon — what agency, or unified sphere of activity, or principle, or lawfulness, or other reality of any sort are they appealing to? They cannot be pointing merely to a particular collection of objects, because the collection can be endlessly varied or perturbed, and yet the context remains more or less coherent, and the organism more or less maintains its character. What is coherent? What has this character?
I would like to conclude by offering a few brief thoughts in little more than aphoristic form — thoughts that I believe will prove essential in finding reasonable answers to questions such as these. I expect to be developing these thoughts further in forthcoming commentaries.
Key thoughts for a biology worthy of life
![]() The problem of form in the organism — how does a single cell
(zygote)
reliably develop to maturity “according to its own kind” — has vexed
biologists for centuries. But the same mystery plays out in the mature
organism, which must continually work to maintain its normal form, as well
as restore it when injured. It is difficult to bring oneself fully face
to face with the enormity of this accomplishment. Scientists can
damage tissues
in endlessly creative ways that the organism has never confronted in its
evolutionary history. Yet, so far as its resources allow, it mobilizes
those resources, sets them in motion, and does what it has never done
before, all in the interest of restoring a dynamic form and a functioning
that the individual molecules and cells certainly cannot be said to
“understand” or “have in view”.
The problem of form in the organism — how does a single cell
(zygote)
reliably develop to maturity “according to its own kind” — has vexed
biologists for centuries. But the same mystery plays out in the mature
organism, which must continually work to maintain its normal form, as well
as restore it when injured. It is difficult to bring oneself fully face
to face with the enormity of this accomplishment. Scientists can
damage tissues
in endlessly creative ways that the organism has never confronted in its
evolutionary history. Yet, so far as its resources allow, it mobilizes
those resources, sets them in motion, and does what it has never done
before, all in the interest of restoring a dynamic form and a functioning
that the individual molecules and cells certainly cannot be said to
“understand” or “have in view”.
We can frame the problem of identity and context with this question: Where do we find the context and activity that, in whatever sense we choose to use the phrase, does “have in view” this restorative aim? Not an easy question. Yet the achievement is repeatedly carried through; an ever-adaptive intelligence comes into play somehow, and all those molecules and cells are quite capable of participating in and being caught up in the play.
![]() It ought to be a truism among biologists that our understanding of
living creatures must be, at its most fundamental (irreducible)
level, an understanding of activity. And therefore our knowledge
of changes in the organism — for example, evolutionary changes, or the
transformations during development — must be, fundamentally, a knowledge
of changes in activity, not mere rearrangements of things with predefined
functional identities. The experimentalist, in comparing two specimens
under different conditions, can note down all the differences in
quantities of this or that molecule, or all the differences in morphology
of leaf or limb. But each of these changes precipitates, so to speak, out
of
an altered process.
It ought to be a truism among biologists that our understanding of
living creatures must be, at its most fundamental (irreducible)
level, an understanding of activity. And therefore our knowledge
of changes in the organism — for example, evolutionary changes, or the
transformations during development — must be, fundamentally, a knowledge
of changes in activity, not mere rearrangements of things with predefined
functional identities. The experimentalist, in comparing two specimens
under different conditions, can note down all the differences in
quantities of this or that molecule, or all the differences in morphology
of leaf or limb. But each of these changes precipitates, so to speak, out
of
an altered process.
The organism is always a becoming, and to think about a capacity for becoming is radically different from thinking about a collection of well-defined (which is to say, context-independent) things. If the biologist’s training included even one week where this difference was held up for attention, everything in biology would be colored differently. We can perhaps glimpse the difficulty of the task by asking: How would we characterize a change in directed (forming) activity, as opposed to a change from one finished form to another, or a mere rearrangement of things?
![]() Our understanding necessarily alternates between narrow focus on the parts
of an activity viewed separately, and receptivity to the larger expressive
picture. The two facets of understanding serve each other. This is
related (rather exactly, I think) to the interplay between our hearing of
particular words as we listen to speech, and our progressive apprehension
of the overall meaning that more and more shines through those words,
modifies their identity, and subordinates them to the developing direction
of thought. We could not understand speech without hearing individual
words, but neither could we understand the individual words in their
current meaning without grasping the overall import of what is being said
(Bortoft 2012*).
Our understanding necessarily alternates between narrow focus on the parts
of an activity viewed separately, and receptivity to the larger expressive
picture. The two facets of understanding serve each other. This is
related (rather exactly, I think) to the interplay between our hearing of
particular words as we listen to speech, and our progressive apprehension
of the overall meaning that more and more shines through those words,
modifies their identity, and subordinates them to the developing direction
of thought. We could not understand speech without hearing individual
words, but neither could we understand the individual words in their
current meaning without grasping the overall import of what is being said
(Bortoft 2012*).
![]() This import, while suffusing and bringing to life the individual sounding
words, is not itself a physical thing that sounds, nor can it be equated
with the sum total of physical sounds. It is, rather, the meaning of the
speech. Similarly, the meaningful agency at work in the organism — the
source of coherent, self-maintaining activity constituting a particular
way of being — cannot be identified with any or all of the physical
processes it brings into movement.
This import, while suffusing and bringing to life the individual sounding
words, is not itself a physical thing that sounds, nor can it be equated
with the sum total of physical sounds. It is, rather, the meaning of the
speech. Similarly, the meaningful agency at work in the organism — the
source of coherent, self-maintaining activity constituting a particular
way of being — cannot be identified with any or all of the physical
processes it brings into movement.
![]() To any for whom this conjoining of meaning/speech and science seems
objectionable: why forget yourselves as living organisms, as if the cosmos
were bereft of you? To discover ourselves in the universe — which is to
say, to discover ourselves as discoverers, or knowers, of the universe —
tells us something about what the universe is like. It does so fully as
much as our discovery of gravitational attraction among objects of the
solar system informs us about the reality of gravity in the universe. If
we find ourselves to be knowers of a knowable world, then knowing and
being known — being cognized, being the stuff of cognition, being
meaningful — belong to the world’s and our own character. Speaking
and being spoken belong to the very fabric of things.
To any for whom this conjoining of meaning/speech and science seems
objectionable: why forget yourselves as living organisms, as if the cosmos
were bereft of you? To discover ourselves in the universe — which is to
say, to discover ourselves as discoverers, or knowers, of the universe —
tells us something about what the universe is like. It does so fully as
much as our discovery of gravitational attraction among objects of the
solar system informs us about the reality of gravity in the universe. If
we find ourselves to be knowers of a knowable world, then knowing and
being known — being cognized, being the stuff of cognition, being
meaningful — belong to the world’s and our own character. Speaking
and being spoken belong to the very fabric of things.
![]() It is not I who force such thoughts upon our attention. Whenever
biologists refer to context, they are invoking the parallel with
speech, or text. Their usage requires at least some effort of
understanding. Organic activity is, in fact, always a special kind of
contextualized expression, and “context” is another word for “that which
has — more or less — the character of a meaningful whole”. Without some
quality of wholeness, it would not be an identifiable context.
It is not I who force such thoughts upon our attention. Whenever
biologists refer to context, they are invoking the parallel with
speech, or text. Their usage requires at least some effort of
understanding. Organic activity is, in fact, always a special kind of
contextualized expression, and “context” is another word for “that which
has — more or less — the character of a meaningful whole”. Without some
quality of wholeness, it would not be an identifiable context.
The word “meaningful” here points, in part, to an ideational content. A collection of arbitrarily designated things — a collection lacking a unity of idea — does not add up to a context. We call a context a “context” because it has a recognizable and integral character possessing some sort of ideal unity. The continual activity of the context (or whole) is the way it remains itself; its becoming is the assertion of its dynamic identity and character (Riegner 2013*; Talbott 2013*).
![]() An example. Twenty-two people randomly milling about in a field do not
make an organic context. But when twenty-two people are arrayed against
each other on a football field, consistently, adaptively, and innovatively
performing according to the aims and choreographic requirements of a game
— then we have a recognizable context. The context cannot be defined
apart from the many ideas making up the aim and the meaning of the game3.
An example. Twenty-two people randomly milling about in a field do not
make an organic context. But when twenty-two people are arrayed against
each other on a football field, consistently, adaptively, and innovatively
performing according to the aims and choreographic requirements of a game
— then we have a recognizable context. The context cannot be defined
apart from the many ideas making up the aim and the meaning of the game3.
![]() “Whole” and “context” are flexible terms. Biologists find wholes within
wholes, and contexts within ever larger contexts. The
cell nucleus
is a context for
replicating
DNA,
but both
DNA
and
nucleus
find their proper place only within the cell as a whole, and the
performance of the cell in turn makes sense only within the context of
tissue, organ, and organism. ... Moreover, as a multicellular organism
develops, cells change their identities (as we saw above), and contexts
interpenetrate, dissolve, and re-form.
“Whole” and “context” are flexible terms. Biologists find wholes within
wholes, and contexts within ever larger contexts. The
cell nucleus
is a context for
replicating
DNA,
but both
DNA
and
nucleus
find their proper place only within the cell as a whole, and the
performance of the cell in turn makes sense only within the context of
tissue, organ, and organism. ... Moreover, as a multicellular organism
develops, cells change their identities (as we saw above), and contexts
interpenetrate, dissolve, and re-form.
![]() All this raises what older readers might be tempted to call the “Alexander
Haig problem”: who is really in charge? It’s a question we might approach
by asking about the relation between the organism and nature as a whole —
or, as we humans are likely to put it, between self and world. How can we
speak of “an organism” when there is no strict dividing line between the
organism and its environment? For example, we have more microbial cells
in our body than cells we call our “own”, and we could not live without
them. Do they belong to us, or to our environment?
All this raises what older readers might be tempted to call the “Alexander
Haig problem”: who is really in charge? It’s a question we might approach
by asking about the relation between the organism and nature as a whole —
or, as we humans are likely to put it, between self and world. How can we
speak of “an organism” when there is no strict dividing line between the
organism and its environment? For example, we have more microbial cells
in our body than cells we call our “own”, and we could not live without
them. Do they belong to us, or to our environment?
Yet we are quite capable of distinguishing where we cannot rigorously divide. Just as the “particle” of the physicist is a center of force interacting with countless other centers of force — yet is still distinguishable as this center — so, too, the organism can be distinguished from its inanimate surround. This doesn’t mean, however, that the line between the individual organism and its ecological/physical context is ever precisely definable. (See the following.)
![]() In our own case, the historian finds a process of individuation at work
from the age of myth to the modern day
(Barfield 1965*).
That is, the tension and separation between self and world increases as
one moves forward along the various advancing fronts of civilization.
This has brought us to a point where, over the past few hundred years —
and particularly in the west — we have found ourselves wrestling with the
problem of isolated and detached subjectivities facing an objective world
that we place (we place) “out there”. Where once we were “at home
in the universe”, experiencing ourselves as microcosms of the macrocosm,
today we are not quite sure how — or whether at all — our individual
selves and the world can be reconciled.
In our own case, the historian finds a process of individuation at work
from the age of myth to the modern day
(Barfield 1965*).
That is, the tension and separation between self and world increases as
one moves forward along the various advancing fronts of civilization.
This has brought us to a point where, over the past few hundred years —
and particularly in the west — we have found ourselves wrestling with the
problem of isolated and detached subjectivities facing an objective world
that we place (we place) “out there”. Where once we were “at home
in the universe”, experiencing ourselves as microcosms of the macrocosm,
today we are not quite sure how — or whether at all — our individual
selves and the world can be reconciled.
![]() It can be argued — it has in fact been argued
(Rosslenbroich 2009*)
— that evolution as a whole likewise shows certain broad thrusts toward
greater individuation:
It can be argued — it has in fact been argued
(Rosslenbroich 2009*)
— that evolution as a whole likewise shows certain broad thrusts toward
greater individuation:
Organisms gained in stability, self-regulation and self-assertion. The direct influences of the environment were gradually reduced and a stabilization of self-referential, intrinsic functions within the systems was generated. In higher animals this included the potential for more flexible and self-determined behaviors. ... Features of an increasing autonomy are spatial separations from the environment, increases in homeostatic functions and in body size, internalizations and an increase in physiological and behavioral flexibility (Rosslenbroich 2011*).
![]() The movement toward individuation results in a changing balance between
organism and environment. This changing balance is reflected in the
entire spectrum of ways we might answer this question: Does the organism
speak, or is it spoken? To what degree does it speak the meaning of its
life from its own center, and to what degree does the world breathe
meaning into and through it?
The movement toward individuation results in a changing balance between
organism and environment. This changing balance is reflected in the
entire spectrum of ways we might answer this question: Does the organism
speak, or is it spoken? To what degree does it speak the meaning of its
life from its own center, and to what degree does the world breathe
meaning into and through it?
Every autumn in North America, migrating monarch butterflies travel southward a couple of thousand miles to a small forest location in Mexico — a location they have never before come near in their lifetime. It seems fair to say in this case that they are moved by the call of their surroundings — surroundings that play into and help define their bodily instincts from without — more than they are moved by any choice of their own. But analogs to choice become more vivid among birds and mammals, and something like true and free choice comes to expression, at least as a potential, in the human being.
![]() The tension between self and other is a tension within unity. The self’s
emergence as a bright focus of self-awareness in the human individual does
not mean that the opposite pole, whether of the larger human society or of
nature in general, must be weakened. (For an inanimate comparison: think
of the bar magnet, with its north and south poles. The stronger the north
pole, the stronger the south pole — and therefore the more profound the
unity of the magnet as a whole, which is capable of holding this greater
tension within itself.)
The tension between self and other is a tension within unity. The self’s
emergence as a bright focus of self-awareness in the human individual does
not mean that the opposite pole, whether of the larger human society or of
nature in general, must be weakened. (For an inanimate comparison: think
of the bar magnet, with its north and south poles. The stronger the north
pole, the stronger the south pole — and therefore the more profound the
unity of the magnet as a whole, which is capable of holding this greater
tension within itself.)
One may be dismayed by the centrifugal and chaotic potentials of the one-sided contemporary obsession with the individual and his rights. But, on the other hand, this is the individual who for the first time, standing firmly on his own ground, can behold the planet as a whole — can feel himself belonging to humankind in a way that was inconceivable in earlier eras, can survey the world of nature and become aware of his own dependence upon it and responsibility for it. The stronger the independent being of the individual becomes, the greater its capacity for reunion with other creatures — now in freely spoken love rather than as the world’s irresistible urging (Solovyov 1985*).
![]() I have been emphasizing that the organism is a becoming. This fact
makes a lie of the overly emphatic conviction that we learn who we are
through a study of evolution. Evolution tells us a great deal — but only
about our past. It doesn’t tell us about the potentials of our becoming
in the present. When we learn that such-and-such a trait of lower animals
is recognizable in ourselves in some form, this knowledge immediately
changes our relation to that trait. It opens up a space of freedom to do
work and act consciously where previously, rather as with the monarch
butterlies migrating south, nature was simply acting through us. “The
truth will make you free”. Those who delight in pointing out our “lower
nature” are actually assisting us — presumably to their great
disappointment, should they become aware of it — toward the realization of
a higher nature.
I have been emphasizing that the organism is a becoming. This fact
makes a lie of the overly emphatic conviction that we learn who we are
through a study of evolution. Evolution tells us a great deal — but only
about our past. It doesn’t tell us about the potentials of our becoming
in the present. When we learn that such-and-such a trait of lower animals
is recognizable in ourselves in some form, this knowledge immediately
changes our relation to that trait. It opens up a space of freedom to do
work and act consciously where previously, rather as with the monarch
butterlies migrating south, nature was simply acting through us. “The
truth will make you free”. Those who delight in pointing out our “lower
nature” are actually assisting us — presumably to their great
disappointment, should they become aware of it — toward the realization of
a higher nature.
![]() We see in the organism’s relation to its environment, then, a unity and
tension between opposites, distantly analogous to the tensive unity of a
magnet possessing north and south poles. This unity varies from what we
see in, say, bacteria — where the distinction between organism and world
is far less pronounced — to the sometimes painful separation from the
world we humans can experience.
We see in the organism’s relation to its environment, then, a unity and
tension between opposites, distantly analogous to the tensive unity of a
magnet possessing north and south poles. This unity varies from what we
see in, say, bacteria — where the distinction between organism and world
is far less pronounced — to the sometimes painful separation from the
world we humans can experience.
Both the tension between opposites and the underlying unity are prerequisites for scientific understanding or any other understanding. We must separate ourselves from what we observe in order to say “I know”, but we must unite ourselves cognitively with what we observe before we can say “I know”.
Modern science has perfected the art of separating itself from the object of observation. It has scarcely begun to acknowledge the unity that is also prerequisite for cognition. The very act of suppressing the subject’s self-awareness in the interest of objectivity helps to sustain what then appears to be an absolute break between subject and object. Nearly all modern philosophy can be viewed as a series of troubled footnotes to this falsely imagined break.
![]()
Notes
1. Doing a quick hyperbole-check on myself, I find that a google scholar search on "context dependent" gene cell yields about 52,000 results. It would be a worthwhile exercise for the reader to execute that search (by clicking here) and then browse down through some of the article titles and excerpts.
2. For other examples of the bizarre juxtaposition of context-dependence and controlling factors, see “Who Regulates the Regulators?” in “The Unbearable Wholeness of Beings”, and also the discussion of circadian rhythms in “From Physical Causes to Organisms of Meaning”.
3. More generally: the almost universal willingness among scientists to ignore the ideational character of the world while passionately devoting themselves to scientific ideas they hope will prove faithful to the world’s reality is one of the more mystifying features of the modern era.
Sources:
Barfield, Owen (1965). Saving the Appearances. New York: Harcourt, Brace and World. Originally published in 1957.
Bortoft, Henri (2012). Taking Appearance Seriously: The Dynamic Way of Seeing in Goethe and European Thought. Edinburgh UK: Floris Books.
Riegner, Mark F. (2013). “Ancestor of the New Archetypal Biology: Goethe’s Dynamic Typology as a Model for Contemporary Evolutionary Developmental Biology,” Studies in History and Philosophy of Biological and Biomedical Sciences. doi:10.1016/j.shpsc.2013.05.019
Rompolas, Panteleimon, Kailin R. Mesa and Valentina Greco (2014). “Spatial Organization within a Niche as a Determinant of Stem-Cell Fate”, Nature vol. 502 (Oct. 24), pp. 513-8. doi:10.1038/nature12602
Rosslenbroich, Bernd (2009). “The Theory of Increasing Autonomy in Evolution: A Proposal for Understanding Macroevolutionary Innovations”, Biology and Philosophy vol. 24, pp. 623-44. doi:10.1007/s10539-009-9167-9
Rosslenbroich, Bernd (2011). “Outline of a Concept for Organismic Systems Biology”, Seminars in Cancer Biology vol. 21, pp. 156-64. doi:10.1016/j.semcancer.2011.06.001
Solovyov, Vladimir (1985). The Meaning of Love, edited with a substantially revised translation by Thomas R. Beyer, Jr., introduction by Owen Barfield. Hudson NY: Lindisfarne Press. Originally published in 1894.
Talbott, Stephen L. (2004). “The Reduction Complex”, NetFuture #158 (Nov. 9). Available at http://BiologyWorthyofLife.org/mqual/ch04.htm.
Talbott, Stephen L. (2013). “Rebirth of the Type: Notes on A Recent Paper by Mark Riegner”, In Context #30 (Fall), pp. 5-6. Available at https://natureinstitute.org/pub/ic/ic30/riegner-on-typology.pdf
Tan, Jiaying (2013). “Stemness in the Niche”, Cell vol. 155 (Dec. 19), pp. 1439-40. doi:10.1016/j.cell.2013.12.003
Further information:
For a brief discussion highlighting the sensitivity of cancer cells to context, see the section, Cancer and Context, in Putting Genes in Perspective.
The relation of part to whole in the organism is also discussed in an article section entitled What Does ‘More Than the Sum of Its Parts’ Mean?.
The dynamic unity discerned in organic processes is discussed in Can We Learn to Think Like a Plant?.
Regarding the “ideational” character of the organism, see in particular From Physical Causes to Organisms of Meaning and How Biologists Lost Sight of the Meaning of Life.
The issues most deeply underlying the thoughts expressed here will never be successfully grasped without an epistemological reorientation — that is, without a changed awareness of our relation to the world as knowers. For two preliminary and somewhat overlapping treatments of this relation, see Reframing the Mind-Body Problem and A Modest Epistemological Exercise.
This document: https://bwo.life/org/comm/ar/2014/who-are-we_15.htm
Steve Talbott :: Who Are You and Who Am I and Who Are We?